Артериальное давление у крокодила
Анатомия и физиология сердечно-сосудистой и дыхательной систем у рептилий. Часть 1
Еще фото
Автор (ы): Л.А. Стоянов, врач ветеринарной медицины, начальник ветеринарно-медицинского отдела экзотических животных Международной ассоциации океанариумов и дельфинариев
Организация(и): Сеть океанариумов «Немо», г. Одесса
Журнал: №1 — 2013
Благодарим редакцию журнал «Мир ветеринарии», Украина, за любезно предоставленную статью Л.А. Стоянова
Анатомия сердечно-сосудистой системы
У рептилий нет одной общей для всех схемы кровообращения. Однако можно выделить два основных типа строения сердца. Первый характерен для чешуйчатых и черепах, а второй – для крокодилов.
Ящерицы, змеи и черепахи
Сердце змей, ящериц и черепах трехкамерное, с двумя предсердиями и одним желудочком (Рис. 1-3). Такое строение предполагает возможность смешивания богатой кислородом крови из легких с кровью, ненасыщенной кислородом, идущей от систем органов. Ряд мышечных гребней и определенная периодичность сокращений служат для функционального разделения желудочка.
Правое предсердие получает ненасыщенную кислородом кровь, которая поступает от всех органов, через венозный синус – расширение на дорсальной стороне предсердия. Стенка венозного синуса мышечная, но не такая толстая, как стенка предсердия. Венозный синус получает кровь от четырех вен:
1. правой передней полой вены;
2. левой передней полой вены;
3. задней полой вены;
4. левой печеночной вены.
Левое предсердие получает насыщенную кислородом кровь из легких через левую и правую легочные вены.
В самом желудочке различают три полости: легочную, венозную и артериальную. Легочная полость – самый вентральный отдел, он продолжается краниально до устья легочной артерии. Артериальная и венозная полости расположены дорсальнее по отношению к легочной и получают кровь от левого и правого предсердия, соответственно. В своей самой краниальной и вентральной части венозная полость дает начало левой и правой дугам аорты (Рис. 4).
Мышечный гребень в некоторой степени отделяет легочную полость от других полостей. Артериальная и венозная полости объединены межжелудочковым каналом.
Одностворчатые атриовентрикулярные клапаны открываются с краниальной стороны межжелудочкового канала. Анатомически они организованы таким образом, что частично закрывают межжелудочковый канал во время систолы предсердия. Во время систолы желудочка их функция заключается в предотвращении регургитации крови из желудочка в предсердия. Серия мышечных сокращений и последующая разница давления в сердце рассматриваемых здесь рептилий разнесены по времени так, чтобы создать функционально двойную систему кровообращения. Систола предсердия нагнетает кровь в желудочек. Расположение атриовентрикулярных клапанов поперек межжелудочкового канала позволяет венозной крови из правого предсердия наполнять венозную и легочную полости. В то же время кровь из легких попадает из левого предсердия в артериальную полость. Желудочковая систола начинается с сокращения венозной полости. Последовательные сокращения венозной и легочной полостей выталкивают кровь из них в малый круг кровообращения с низким давлением.
В продолжение систолы сокращается артериальная полость, что приводит к движению крови через частично сокращенную венозную полость в большой круг кровообращения через левую и правую дуги аорты. Сокращение желудочка приводит мышечный гребень в положение непосредственной близости к вентральной стенке желудочка, создавая таким образом перегородку между артериальной и легочной полостями. Левый и правый атриовентрикулярные клапаны предотвращают возврат крови из желудочка в предсердия.
Все вышеупомянутые явления протекают только при нормальном дыхании. Такая система подачи крови ведет к ее сбросу слева направо на основе разницы давлений. При погружении под воду или в других ситуациях, когда легочное сопротивление и давление повышаются, движение крови происходит справа налево. У красноухой черепахи (Trachemys scripta elegans) при нормальном дыхании кровь движется преимущественно в легочном круге, который получает 60% объема выходящей из сердца крови, а оставшиеся 40% направляются ко всем системам органов. При погружении в воду кровь преимущественно движется по большому кругу, минуя легкие. В таких обстоятельствах давление в легочных сосудах выше, чем на периферии, поэтому кровь входит в сосуды с меньшим давлением – в дуги аорты. У ящериц кровь проходит в основном по левой дуге.
Крокодилы
Строение сердца у крокодилов очень напоминает таковое у птиц и млекопитающих, с той лишь разницей, что у крокодилов есть небольшое отверстие в межжелудочковой перегородке, разделяющей правый и левый желудочки – паниццево отверстие (foramen Pannizi), и что левая дуга аорты выходит из правого желудочка.
Строение сердца крокодилов двойственно по своей природе. Некоторое смешивание насыщенной и ненасыщенной кислородом крови может происходить через паниццево отверстие или в спинной аорте в месте слияния правой и левой дуг. Впрочем, при нормальном дыхании последнего варианта смешивания не происходит, так как давление в системном круге кровообращения превосходит давление в легочном круге. Сброс крови слева направо происходит через паниццево отверстие, и небольшое количество насыщенной кислородом крови попадает в правый желудочек.
Во время погружения под воду или в других условиях, при которых повышается сопротивление легочных сосудов, давление в легочной артерии также существенно возрастает. В результате кровь отводится от легких в системный круг кровообращения. Таким образом, кровь поступает преимущественно в левую дугу аорты, а не в легочную артерию. Существует мнение, что причиной возникновения высокого легочного сопротивления при погружении и, как следствие, сброса крови справа налево, является особый путь оттока крови через правый желудочек. В нем есть отдельная «камера», субпульмонарный конус, который благодаря задержке деполяризации и зубчатым клапанам контролирует поступление крови в легочную сосудистую сеть.
Сам факт сброса крови справа налево при задержке дыхания и повышении сопротивления легочных сосудов может иметь большое клиническое значение. Рептилии под наркозом или без дыхания в отсутствии искусственной вентиляции легких могут демонстрировать непрогнозируемые реакции на ингаляционную анестезию. Кровообращение в обход легких может приводить к недостаточному распределению анестезирующих газов, таких как изофлюран, в системном круге для дальнейших манипуляций под анестезией. Значение длительного сброса справа налево, который может отмечаться при хронических воспалительных процессах в легких, до сих пор мало изучено. При этом можно ожидать серьезных изменений со стороны сердечно-сосудистой системы.
Воротная система почек
Воротная система почек – одна из частей венозной системы рептилий, вызывающая множество вопросов, потенциально имеющих значение для врача. Ее функция заключается в обеспечении достаточного кровоснабжения почечных канальцев при замедлении тока крови через клубочки для сохранения воды.
Афферентные вены воротной системы почек не проникают в клубочки; вместо этого они снабжают кровью проксимальные и дистальные извитые канальцы. Как и у млекопитающих, кровь к клеткам канальцев у рептилий подают приносящие артериолы, которые выходят из клубочков. Однако, в отличие от млекопитающих, в нефронах рептилий нет петель Генле и, следовательно, не происходит реабсорбции воды. В результате, для того чтобы сохранить воду, под воздействием аргининвазо-тоцина замедляется приносящий ток крови через клубочки. При снижении кровоснабжения в клубочках воротная система почек жизненно необходима для подачи крови к канальцам во избежание циркуляторного некроза.
Физиология сердечно-сосудистой системы
Частота сердцебиений у рептилий находится в довольно сложной зависимости от ряда факторов, в том числе температуры тела, размеров тела, уровня обмена веществ, дыхания и внешних раздражителей. Сердечная мышца характеризуется присущей ей максимальной производительностью, измеряемой максимальным напряжением сокращения в пределах зоны оптимальных предпочитаемых температур (ЗОПТ) для данного вида. В общем случае повышение активности приводит к повышению частоты сердцебиений. Частота может увеличиваться втрое по сравнению с частотой сокращений в состоянии покоя. Также, как правило, существует обратная зависимость между размером тела и частотой сердцебиений при заданной температуре.
Интересные вариации частоты сердцебиений при одной и той же температуре окружающей среды проявляются в зависимости от температурного статуса рептилии. В процессе нагревания животное обычно имеет более высокую частоту сердцебиений, чем во время охлаждения. Ускорение сердечного ритма при прогреве помогает достигнуть максимального поглощения тепла. Снижение частоты ударов сердца при понижении температуры окружающей среды помогает рептилии замедлить потерю тепла.
При низких температурах минутный объем сердца, по-видимому, поддерживается за счет увеличения его ударного объема. Ускоренное сердцебиение при повышенных температурах, очевидно, связано со скоростью обмена веществ. Теоретически, высокая частота сердцебиений должна ускорять транспорт кислорода. Изучение кислородного пульса (количество потребляемого кислорода при каждом сердцебиении в мл на массу тела в г) у разных видов говорит об отсутствии последовательной схемы взаимосвязи между объемом сокращения, поглощением кислорода и частотой сердцебиений ввиду повышенной потребности в кислороде при увеличении скорости метаболизма. Различные виды рептилий предположительно обладают множеством механизмов для улучшения подачи кислорода во время ускорения обмена веществ. Отдельно следует упомянуть тот факт, что скорость сердцебиений стремится к увеличению во время активного дыхания и уменьшению при задержке дыхания. Увеличение сердечного ритма совпадает со снижением легочного сопротивления и последующим увеличением легочного кровообращения. Соответственно, увеличение легочного кровообращения в период увеличения дыхательной активности служит для большей эффективности газообмена.
Сердечно-сосудистая система играет ключевую роль в терморегуляции рептилий. Как уже было сказано, частота ударов сердца повышается, когда животное нагревается, и понижается при его охлаждении. Несмотря на то что контролирующий механизм до конца не известен, изменения в кровеносной системе происходят до того, как изменится общая температура тела, что предполагает наличие кожных терморецепторов и барорецепторов.
При нагревании кожных покровов отмечается расширение сосудов в коже. Отток крови в периферические сосуды приводит к падению общего кровяного давления. Понижение сопротивления периферических сосудов способствует развитию сброса крови в сердце справа налево. Кровяное давление поддерживается, таким образом, на уровне, достаточном для снабжения кровью мозга и органов чувств по правой дуге аорты. Кроме того, так как кровь из кожных покровов возвращается в общее кровяное русло, повышается общая температура тела.
Снижение частоты сердцебиений при охлаждении кожи служит для сохранения тепла. При этом наблюдается сужение кровеносных сосудов в коже и относительное расширение сосудов в мышцах. Такое перераспределение крови призвано замедлить теплоотдачу.
Так же как для птиц и млекопитающих, изменения гемодинамики при погружении под воду очень важны и для рептилий. У них имеется ряд преимуществ по сравнению с теплокровными животными, так как рептилии могут использовать альтернативный путь метаболизма при отсутствии кислорода – анаэробный гликолиз. Способность выдерживать анаэробиоз различна у разных видов рептилий. Некоторые ящерицы выдерживают без кислорода не более 25 минут, тогда как некоторые виды черепах способны задерживать дыхание на 33 часа и более. Основные различия заключаются в разной толерантности миокарда к гипоксии.
Как правило, при погружении под воду развивается брадикардия. У крокодилов она обусловлена вагальным торможением сердца под некоторым влиянием торакального или внутрилегочного давления. При нырянии происходит симпатическое сужение кровеносных сосудов в скелетных мышцах, часто до ишемического порога. Такое повышение периферического сопротивления поддерживает кровяное давление для нормальной работы органов.
Сброс крови справа налево возникает при истощении запаса кислорода в паренхиме легких. При дальнейшем погружении сброс справа налево доминирует, практически полностью исключая подачу крови к легким. Общий сердечный выброс может снижаться до уровня в 5% по сравнению с нормальным состоянием. Способность снижать до минимума рабочую нагрузку на сердце, прокачивая лишь небольшую часть крови в системное русло, обеспечивает рептилиям явное преимущество при погружениях по сравнению с птицами и млекопитающими. Брадикардия, связанная с погружением, быстро обратима при первом же вдохе; у некоторых видов даже отмечено ускорение работы сердца еще до выхода на сушу.
Свойства кровеносной системы и их связь с газообменом на клеточном уровне должны приниматься во внимание в любых исследованиях в области кардиологии рептилий. Несмотря на кажущуюся несущественность данного вопроса, клинически подтверждено, что изменения в функционировании сердца или легких могут в значительной степени влиять на способность кровеносной системы к переносу кислорода и углекислого газа.
Молекула гемоглобина считается компонентом, от которого зависят респираторные свойства крови. Хотя структура гемоглобина рептилий пока полностью не описана, она, скорее всего, такая же, как у других позвоночных. Тем не менее, известен ряд существенных отличий в способности гемоглобина удерживать и отдавать кислород. Для этих отличий не было найдено каких-либо закономерностей в зависимости от условий среды, и они не являются общими для всего класса рептилий.
В целом, сродство крови к кислороду зависит от вида рептилии, возраста, размеров и температуры тела. Количество кислорода в организме животного определяется гематокритом и объемом крови. Способность крови переносить кислород зависит от количества эритроцитов на единицу объема (гематокрита). У рептилий он варьирует в пределах: 5-11% у черепах, 6-15% у крокодилов, 8-12% у змей, и от 7% до 8% у ящериц.
По мере растворения кислорода его давление (мера концентрации) приводит к насыщению или частичному насыщению гемоглобина. Молекула гемоглобина отвечает за респираторные свойства и цвет крови. Кривые диссоциации кислорода показывают, какое его количество удерживается гемоглобином при определенных условиях, и отражают влияние температуры, рН, углекислого газа, продуктов гликолиза, органических фосфатов в эритроцитах и таких ионов, как Na+, K+, Mg2+, Cl — , SO4 2- .
Если гемоглобин претерпевает изменения с момента рождения до формирования взрослой особи, то способность крови к насыщению кислородом будет различной в зависимости от этапа онтогенетического развития. При высокой скорости обмена веществ кривые диссоциации кислорода будут смещаться вправо, то есть сродство крови к кислороду будет ниже, что упрощает его доставку к тканям. У рептилий кривые диссоциации кислорода крайне вариабельны. Их сложно обобщить в связи с влиянием непостоянной температуры и скорости метаболизма, а также других перечисленных ранее факторов.
Разные рептилии обладают разными формами гемоглобина, и у некоторых видов гемоглобин эмбриона может иметь сходство к кислороду, отличное от такового у взрослых особей. Гемоглобин может по-разному принимать и отдавать кислород. Эти отличия часто не обнаруживаются клинически, но о них необходимо помнить, чтобы избежать излишней экстраполяции с одного вида на другой.
Сродство к кислороду является мерой того, насколько легко гемоглобин отдает кислород тканям. Гемоглобин с высоким сродством отдает кислород хуже. Низкое сродство означает лучшую отдачу кислорода. У рептилий обычно сродство гемоглобина к кислороду ниже, чем у млекопитающих. Эта адаптация позволяет снабжать кислородом ткани даже при небольшом его содержании в крови.
Во время нагрузок или стресса рептилии могут испытывать метаболический ацидоз вследствие образования молочной кислоты. Изменение рН крови снижает ее сродство к кислороду (эффект Бора), что приводит к тому, что кровь удерживает меньше кислорода и быстрее отдает его тканям.
Изучение кривых диссоциации кислорода у ряда видов рептилий не выявило для них определенных закономерностей. Однако можно предложить несколько общих концепций для отдельных групп рептилий.
Среди ящериц самые активные виды (например, те-йиды, веретеницевые) обладают, как и следовало ожидать, более низким сродством к кислороду. Более высокое сродство к кислороду характерно для медлительных рептилий или для хищников, поджидающих свою добычу (например, хамелеоны, гекконы). Некой серединой для сравнения можно считать игуановых (в том числе, Iguana iguana, Anolis spp., Ctenosaura spp.). Известно, что у игуановых ящериц сродство крови к кислороду напрямую связано с размерами тела. Однако данные, полученные путем измерений при предпочитаемой температуре, слишком ненадежны ввиду поведенческих различий между видами и потому не могут считаться клинически значимыми.
У черепах видимая разница существует между водными и сухопутными видами. Как правило, у водных видов сродство к кислороду ниже, то есть отдача кислорода происходит лучше. У некоторых черепах, живущих в условиях постоянной гипоксии, кровь обладает буферными свойствами, задерживающими эффект Бора, что можно считать адаптацией, связанной с необходимостью максимальной отдачи кислорода во время погружения. Неожиданным исключением является иловая красноватая черепаха (Kinosternum subrubrum), у которой кривая диссоциации кислорода такая же, как и у наземных черепах.
Змеи в этом вопросе принципиально отличаются от черепах. Сравнение водяной яванской бородавчатой змеи (Acrochordus javanicus) и обыкновенного удава (Constrictor constrictor) показало их противоположность по сродству к кислороду. У водяной змеи сродство к кислороду было выше, чем у наземной.
Эта разница может отчасти быть результатом усиленного эффекта Бора, отмечаемого у водных змей. Роль увеличения эффекта Бора, по-видимому, заключается в том, чтобы обеспечить доступность большего количества кислорода в периоды без дыхания при возрастании уровня CO2 в крови. Такая система насыщения крови кислородом позволяет этим видам отдавать кислород, когда это необходимо, во время погружения, и принимать кислород, когда он наиболее доступен, во время дыхательной вентиляции. У змей сродство к кислороду снижается с возрастом, тогда как кислородная емкость (процентный объем кислорода в полностью насыщенной крови) увеличивается по мере роста. Влияние размера тела на сродство к кислороду неодинаково; оно снижается с увеличением размеров (с возрастом) у змей, но повышается у ящериц.
Как и следовало ожидать, кислородная емкость достигает максимума, когда рептилия находится в зоне оптимальных предпочитаемых температур. У змей в связи с нерегулярным типом питания сродство к кислороду падает и его потребление резко возрастает во время переваривания пищи (процесса, требующего усиления обмена веществ). После приема большого количества пищи увеличивается не только потребление кислорода, но и размеры сердца. Андерсон и др. отмечают, что скорость обмена веществ после еды у тигрового питона (Python molurus bivitattus) может увеличиваться до 40%. Высокий уровень метаболизма может сохраняться до 14 дней.
Для поддержания такого уровня обмена сердце питона гипертрофируется на протяжении 48 часов после употребления пищи. Масса сердца может увеличиваться на 40% в ответ на увеличение экспрессии генов сократительных белков мышц. После завершения переваривания пищи размеры сердца возвращаются к норме.
Окончание статьи в следующем номере журнала.
НАУКА: Сердце крокодила
НАУКА: Сердце крокодила
Автор: Дмитрий Шабанов
Расскажу историю, которая произошла несколько лет назад. Сейчас я пишу школьный учебник зоологии по программе, в составлении которой участвовал сам. Когда эта версия программы только задумывалась, я убеждал министерского работника [Не российского министерства, не волнуйтесь!], что перед систематическим изучением отдельных групп нужно рассмотреть достаточно большую тему, где будет рассказано о животных вообще.
«Хорошо, а с чего ее начать?» — спросил меня чиновник. Я сказал, что образ жизни животных определяется в первую очередь тем, что они едят и как передвигаются. Значит, надо начинать с разнообразия способов питания. «Вы что! — воскликнул мой собеседник. — Как я понесу такую программу министру? Он сразу спросит, зачем мы внушаем детям, что самое главное — это жрачка!»
Я попытался спорить. Вообще, разделение живых организмов на царства (животные, растения, грибы и прочие) связано прежде всего со способом питания, который, в свою очередь, определяет особенности их строения. Особенности многоклеточных животных — следствие того, что они нуждаются во внешних источниках органических веществ и при этом не впитывают их через поверхность тела, а поедают кусками. Животные — это существа, поедающие другие организмы или их части! Увы, мой собеседник был непреклонен. Министра будет в первую очередь интересовать воспитательный аспект программы.
Размышляя о том, как иначе организовать вводную часть, я сделал тогда непростительную ошибку. Следующей моей идеей стало предложение начинать изучение курса зоологии с разнообразия жизненных циклов. Когда мой собеседник понял, что в качестве «главного в жизни» я собираюсь рассматривать не еду, а размножение, он, кажется, решил, что я над ним издеваюсь… В конце концов я написал что-то, что, как я надеялся, никого не будет шокировать. Потом над этой программой поколдовали методисты, которые исправили в ней все, чего не понимали, и заменили формулировки такими, которые были в ходу в исторические эпохи, когда эти самые методисты учились в пединститутах. Потом злосчастную программу подправили чиновники, потом ее переосмыслили в духе новых руководящих указаний, потом… — в общем, пишу учебник по «собственной» программе и не устаю чертыхаться.
А вспомнил я эту печальную историю потому, что еще раз убедился: для животных таки самое главное — пресловутая «жрачка». Сравнивая разные группы наших родственников друг с другом, мы часто не осознаем, какие особенности привели их к успеху или неудаче. Знаете, например, что стало одним из главных козырей млекопитающих? Успевающий школьник назовет выкармливание потомства молоком, теплокровность, высокое развитие нервной системы или какое-нибудь еще свойство, которое стало возможным благодаря достаточному количеству энергии, получаемой с пищей. А один из главных козырей млекопитающих — строение челюстей и зубов!
Попробуйте пошевелить своей нижней челюстью: вверх-вниз, вправо-влево, вперед-назад. Ее «подвеска» допускает движение во всех трех плоскостях! К тому же на челюстях млекопитающих сидят зубы, строение которых определяется той задачей, что им отведена, — пронзать, давить, перетирать, резать, дробить, откусывать, рвать, удерживать, грызть, мять, поддевать, размалывать, скоблить, etc. Наши челюсти — эволюционный биомеханический шедевр. Кроме млекопитающих, почти никакие наземные позвоночные не способны откусывать от пищи куски! К немногим исключениям принадлежит архаичная гаттерия, способная отпилить челюстями голову птенцу буревестника, и черепахи, отказавшиеся от зубов в пользу рогового ножницеобразного клюва. И хищные птицы, и крокодилы не откусывают куски пищи, а попросту отрывают — упираясь когтями (первые) или крутясь всем телом (вторые).
Кстати, о крокодилах — эта колонка посвящена прежде всего им. Благодаря изощренным экспериментам биологов из Университета Юты удалось узнать кое-что новое о функционировании сердца этих пресмыкающихся. Но прежде — еще несколько слов о школьной биологии.
Некоторые особенности подачи биологического материала сохранились с тех времен, когда школа должна была формировать материалистическое мировоззрение, пропагандируя эволюцию. Вообще говоря, факт эволюции не имеет особого отношения к дилемме «материализм-идеализм» (отказываясь на словах от замшелого диамата, мы почему-то до сих пор придаем чрезмерное значение этой сомнительной дихотомии). Увы, когда вместо современных представлений об эволюции преподаются какие-то лежалые догмы, это только наносит естественнонаучному мировоззрению ущерб. К числу таких догм относится линейное представление об эволюции. Подумайте, история позвоночных — это «куст» из многих ветвей, каждая из которых шла своим путем, приспосабливалась к своему образу жизни. А школьный учитель, перепрыгивая с ветки на ветку этого куста, строит прогрессивную последовательность из «типичных представителей»: ланцетник-окунь-лягушка-ящерица-голубь-соба-ка. Но лягушка никогда не пыталась стать ящерицей, она живет своей жизнью, и без учета этой жизни (и предыстории лягушек) понять ее невозможно!

Что расскажет школьный учитель о крокодилах? Он использует их для иллюстрации утверждения, что наипрогрессивнейшими являются животные с четырехкамерным сердцем и «теплокровностью» (гомойотермные). И — смотрите, дети! — у крокодила четырехкамерное сердце, почти-почти как у млекопитающих и птиц, вот только осталось одно лишнее отверстие. Мы своими глазами видим, как крокодил хотел стать человеком, но не дошел, остановился на полпути.
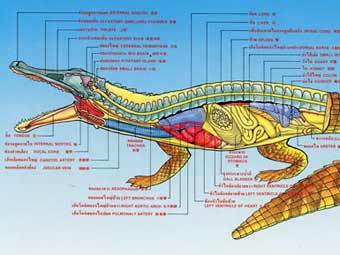
Итак, крокодил имеет четырехкамерное сердце. Из правой его половины кровь идет к легким, из левой — к большому кругу кровообращения (к органам-потребителям полученного в легких кислорода). Но между основаниями отходящих от сердца сосудов находится брешь — паницциево отверстие. В нормальном режиме работы сердца часть артериальной крови переходит через это отверстие из левой половины сердца в правую половину и попадает в левую дугу аорты (посмотрите на рисунок, чтоб не запутаться в право-левых отношениях!). От левой дуги аорты отходят сосуды, идущие к желудку. От левого желудочка отходит правая дуга аорты, питающая голову и передние конечности. А затем дуги аорты сливаются в спинную аорту, которая и обеспечивает кровоснабжение всего остального тела. Почему так сложно?
Для начала разберемся, зачем вообще нужно два круга кровообращения. Рыбы обходятся и одним: сердце — жабры — органы-потребители — сердце. Тут ответ ясен. Легкие не выдержат того давления, которое нужно, чтобы прокачать кровь через все тело. Именно поэтому правая (легочная) половина сердца слабее левой; потому-то нам и кажется, что сердце расположено в левой части грудной полости. Но почему часть крови, текущей по большому кругу кровообращения (от левой половины сердца), проходит у крокодилов через правую, «легочную» часть сердца и левую дугу аорты? У человека неполное разделение потоков крови может вызываться пороком сердца. Зачем такой «порок» крокодилам? Дело в том, что сердце крокодила — не недоделанное сердце человека, оно «задумано» сложнее и может функционировать в двух разных режимах! Когда крокодил активен, обе дуги аорты несут артериальную кровь. Но если паницциево отверстие закрыть (а крокодилы «умеют» это делать), в левую дугу аорты пойдет венозная кровь.
Традиционно такое устройство объясняют тем, что оно якобы позволяет затаившемуся на дне крокодилу отключать легочное кровообращение. Венозная кровь при этом отправляется не в легкие (которые все равно невозможно вентилировать), а сразу в большой круг — по правой дуге аорты. В голову же и к передним ногам пойдет несколько «лучшая» кровь, чем к другим органам. Но если легкие отключены, много ли толку гонять кровь по кругу?
Американские биологи придумали, как проверить давнее предположение о том, что крокодилы перебрасывают кровь из одного круга кровообращения в другой не ради того, чтобы затаиться, а ради лучшего переваривания пищи (углекислый газ — субстрат для производства кислоты железами желудка). Исследователи убедились, что у здоровых молодых аллигаторов в процессе переваривания пищи по левой дуге аорты (той, которая снабжает кровью пищеварительную систему) течет венозная, обогащенная углекислотой кровь. Затем они начали вмешиваться в работу сердца подопытных крокодилов хирургическими методами. У некоторых из них принудительно блокировалась переброска венозной крови в левую дугу аорты; с другими проводилась операция, имитирующая такое вмешательство. Эффект оценивался путем измерения активности желудочной секреции и путем рентгеновского наблюдения за перевариванием проглоченных крокодилами бычьих позвонков. Кроме того, в несчастных аллигаторов помещали полупроводниковые сенсоры, позволявшие измерять их температуру тела. В результате этих манипуляций удалось убедительно подтвердить выдвинутую гипотезу — переброс венозной крови в большой круг кровообращения усиливает выработку кислоты в желудке и ускоряет переваривание пищи.
Крокодилы способны питаться достаточно крупными жертвами, заглатывая добычу целиком или большими кусками (помните, что мы говорили об устройстве челюстей?). Температура тела этих хищников непостоянна, и если они не успеют переварить добычу достаточно быстро, они попросту отравятся ею. Усложненное строение кровеносной системы и ее способность работать в двух разных режимах — способ активизировать переваривание. И свое предназначение пищеварительная система крокодилов оправдывает: на сериях рентгеновских снимков видно, как в желудках хищников «тают» в кислоте солидные бычьи позвонки!
Итак, теперь мы знаем, что главное в жизни крокодилов. Какие цельные все-таки существа!
Крокодилы используют венозную кровь для пищеварения
Исследователи из Университета Чикаго объяснили особенности строения кровеносной системы крокодилов. В экспериментах с американскими аллигаторами им удалось показать, что возможность пускать венозную кровь в обход легких к тканям тела необходима им для переваривания пищи. Работа ученых опубликована в журнале Physiological and Biochemical Zoology.
У крокодилов, как и у других рептилий, сохранились правая и левая дуги аорты. Однако, в отличие от остальных пресмыкающихся, сердце крокодила четырехкамерное, то есть, разделяется на два предсердия и два желудочка.
От левого желудочка отходит правая дуга аорты, по которой насыщенная кислородом после циркуляции через легкие кровь идет в ткани и органы. Левая дуга аорты отходит от правого желудочка и несет венозную кровь, содержащую мало кислорода. На выходе из сердца происходит частичное смешение венозной и артериальной крови из двух дуг аорты. Смешение венозной и артериальной крови характерно для несовершенных кровеносных систем амфибий и рептилий.
Однако крокодилы могут «перекрыть» соединение между дугами аорты. В этом случае венозная кровь из левой дуги не смешивается с артериальной кровью из правой. То есть, основное кровообращение протекает по схеме, характерной для млекопитающих.
Левая дуга аорты ведет к желудку крокодила. При «перекрывании» соединения дуг венозная кровь, текущая по левой дуге, попадает прямо туда. Ученым удалось показать, что в находящихся в желудке железах происходят реакции с участием углекислого газа крови, в результате которых образуются бикарбонат и кислота, которая помогает крокодилу переваривать кости своих жертв. Концентрация кислоты в желудке крокодила при активном пищеварении более чем в десять раз превышает концентрацию, характерную для млекопитающих.
Крокодилы известны тем, что способны переваривать огромные количества пищи — до четверти их собственного веса. Если искусственно помешать венозной крови попадать в желудок в обход легких, пищеварение крокодила нарушается, и он не справляется с перевариванием своей обычной пищи.
Ученые выдвигают несколько предположений, объясняющих такую высокую концентрацию кислоты. Во-первых, кислота препятствует размножению бактерий, что особенно актуально, если учесть, что недопереваренная еда находится в желудке крокодила достаточно долгое время. Во-вторых, бикарбонат необходим крокодилам для нейтрализации большого количества молочной кислоты, которая образуется в мышцах при нападении на жертву. Если вовремя не «очистить» кровь, доза молочной кислоты может оказаться смертельной. «Запасной путь» помогает крокодилам сделать это.
В качестве третьей возможной причины ученые называют необходимость быстро секретировать большое количество кислоты. Это особенно важно для юных крокодилов. Пищеварение лучше протекает в тепле, а теплые места привлекательны также и для естественных врагов, которых много у не вступившего в полную силу молодняка. Как только крокодил попадает в тепло, он должен начать переваривать пищу, и для этого ему необходимо быстро секретировать много кислоты, для чего он использует «перекрывание» дуг аорты.
Источники: http://vetpharma.org/articles/106/4906/, http://document.wikireading.ru/38145, http://lenta.ru/news/2008/02/05/digest/

